Пропионовокислое брожение
Из рассмотренных двух типов брожения видно, что ключевым соединением в обоих процессах является пируват, поскольку в конечном итоге специфика брожения определяется дальнейшей судьбой пирувата. Основная задача последующих реакций — регенерирование молекулы НАД+ и возвращение ее в клеточный метаболизм. Прямое восстановление пирувата с помощью НАД-H2 до молочной кислоты реализуется в молочнокислом брожении. Другая возможность регенерирования НАД+ — "сбрасывание" водорода с НАД-H2 на фрагменты, образуемые при метаболизме пирувата, — имеет место в спиртовом брожении, осуществляемом дрожжами и некоторыми видами бактерий. Третья возможность связана с синтетическим процессом — усложнением молекулы пирувата, в результате которого создается более окисленная молекула акцептора, способная принять больше электронов с восстановленных переносчиков. Это происходит при присоединении к молекуле пирувата CO2, приводящем к формированию четырехуглеродного скелета. Процесс получил название гетеротрофной ассимиляции углекислоты.
|
|
|
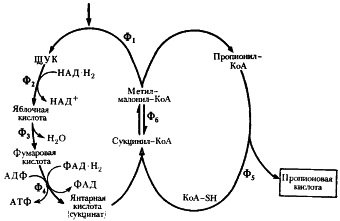
Рис. 54. Превращение пировиноградной кислоты в пропионовую при пропионовокислом брожении: Ф1 — метилмалонил-КоА-карбоксилтрансфераза; Ф2 — малатдегидрогеназа; Ф3 — фумараза: Ф4 — фумаратредуктаза; Ф5 — КоА-трансфераза; Ф6 — метилмалонил-КоА-мутаза (по Daglev, Nicholson. 1973; Rose. 1971) |
Впервые гетеротрофная ассимиляция углекислоты была обнаружена в 1936 г. X. Вудом и К. Веркманом (Н. Wood, C. Werkman) при изучении сбраживания глицерина пропионовыми бактериями. Карбоксилирование пирувата, приводящее к образованию щавелевоуксусной кислоты, получило название реакции Вуда — Веркмана. У эубактерий обнаружены различные реакции карбоксилирования пирувата или его фосфорилированного производного. Показано, что реакции карбоксилирования имеют место у всех гетеротрофных прокариот, а также в клетках всех эукариотных организмов, включая высшие растения и животных. Кроме того, в больших масштабах в природе реакции связывания CO2 осуществляются автотрофными. организмами в процессе хемо- и фотосинтеза.
В пропионовокислом брожении мы имеем дело с реализацией третьей возможности превращения пирувата — его карбоксилированием, приводящим к возникновению нового акцептора водорода — ЩУК. Восстановление пировиноградной кислоты в пропионовую у пропионовокислых бактерий протекает следующим образом (рис. 54). Пировиноградная кислота карбоксилируется в реакции, катализируемой биотинзависимым ферментом, у которого биотин выполняет функцию переносчика CO2. Донором CO2-группы служит метилмалонил-КоА. В результате реакции транскарбоксилирования образуются ЩУК и пропионил-КоА:
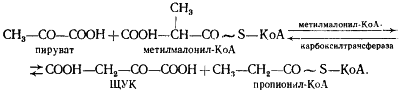
Рассмотрим теперь дальнейшую судьбу каждого из двух продуктов реакции, а также вопрос о происхождении одного из субстратов реакции — метилмалонил-КоА. (Основным источником пировиноградной кислоты служит процесс гликолитического расщепления гексоз или окислительные превращения, если в качестве субстрата брожения используют, например, диоксиацетон или глицерин.)
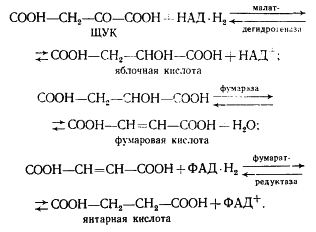
ЩУК в результате трех ферментативных этапов (аналогичных реакциям 6, 7, 8 цикла трикарбоновых кислот, см. рис. 92) превращается в янтарную кислоту:
Следующая реакция заключается в переносе КоА-группы с пропионил-КоА на янтарную кислоту (сукцинат), в результате чего образуется сукцинил-КоА и пропионовая кислота:
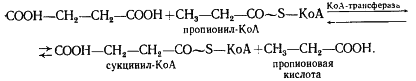
Образовавшаяся пропионовая кислота выводится из процесса и накапливается вне клетки. Сукцинил-КоА превращается в метилмалонил-КоА:
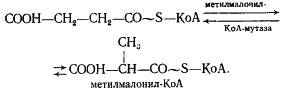
В состав кофермента метилмалонил-КоА-мутазы входит витамин B12. Перегруппировки типа, указанного в приведенном выше уравнении, характерны для реакций, катализируемых ферментами, содержащими витамин B12. В описанной выше реакции происходящие перемещения атомов в молекуле сводятся к двум типам: изменению углерод-углеродных связей и перераспределению водорода между углеродными атомами (рис. 55). Реакция, катализируемая мутазой, — ключевая в пропионовокислом брожении, так как в ней подготавливается субстрат, являющийся предшественником пропионовой кислоты.
Дополнительно
Лазерная медицинская установка для целей лучевой терапии Импульс-1
В настоящее
время лазерное излучение с большим или меньшим успехом применяется в различных
областях науки. Уникальные свойства излучения лазеров, такие, как монохроматичность,
когерентность, малая расходимость и возможность при фокусировке получать очень
высокую плотность мощности на облучаемой по ...
Взаимозаменяемость, стандартиризация и технические измерения
Выполнение
данной курсовой работы преследует собой следующие цели:
–
научить студента самостоятельно применять полученное знание по курсу ВСТИ на
практике;
–
изучение методов и процесса работы со справочной литературой и информацией
ГОСТ;
–
приобретение необхо ...