Бактериородопсин для хранения данных
Молекулярная электроника определяется как кодирование (запись), обработка и распознавание (считывание) информации на молекулярном и макромолекулярном уровне. Основное преимущество молекулярного приближения заключается в возможности молекулярного дизайна и производства приборов "снизу вверх", т.е. атом за атомом или фрагмент за фрагментом, - параметры приборов определяются органическим синтезом и методами генной инженерии. Двумя общепризнанными достоинствами молекулярной электроники являются значительное уменьшение размеров устройств и времени срабатывания (gate propagation delays) логических элементов. В принципе, темпы развития компьютерных технологий таковы, что размеры полупроводниковых чипов согласно закону Мура приблизятся к молекулярным аналогам около 2030 года, что, однако, потребует значительных финансовых затрат. Выходом из положения может стать гибридная молекулярная и полупроводниковая технология, первым коммерческим успехом которой стали жидкокристаллические дисплеи (LCD). Биоэлектроника, являющая разделом молекулярной электроники, исследует возможность применения биополимеров в качестве управляемых светом или электрическими импульсами модулей компьютерных и оптических систем (Birge R.R., 1999). Основное требование к вероятным кандидатам среди большого семейства биополимеров состоит в том, что они должны обратимо изменять свою структуру в ответ на некое физическое воздействие и генерировать, по крайней мере, два дискретных состояния, отличающихся легко измеряемыми физическими характеристиками (например, спектральными параметрами). Значительный интерес в связи с этим представляют белки, основная функция которых связана с трансформацией энергии света в химическую в различных фотосинтетических системах. Наиболее вероятным кандидатом среди них является светозависимый протонный насос - бактериородопсин (БР) из галофильного микроорганизма Halobacterium salinarum (ранее Halobacterium halobium), открытый в 1971 году (Oesterhelt D., Stoeckenius W., 1971). Бактериородопсин - ретиналь-содержащий генератор протонного транспорта представляет собой трансмембранный белок в 248 аминокислот с молекулярным весом 26 кДа, пронизывающий мембрану в виде семи a-спиралей; N- и C-концы полипептидной цепи находятся по разные стороны цитоплазматической мембраны: N-конец обращен наружу, а C-конец - внутрь клетки (рис.1, 2).
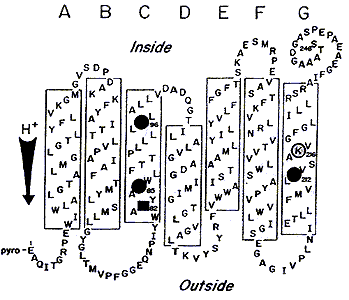 Рис.1.
Рис.1.
Модель БР в элементах вторичной структуры. Выделены аминокислоты, участвующие в протонном транспорте: кружками остатки аспарагиновой кислоты, квадратом остаток аргинина. С Lys-216 (К-216) образуется основание Шиффа (SB).Стрелкой показано направление протонного транспорта.
Хромофор БР - протонированный альдимин ретиналя с e - аминогруппой остатка Lys-216 размещен в гидрофобной части молекулы. После поглощения кванта света в ходе фотоцикла происходит изомеризация ретиналя из all-E в 13Z-форму. Белковое микроокружение хромофора может рассматриваться как рецептор с субстратной специфичностью для all-E /13Z-ретиналя, который катализирует эту изомеризацию при комнатной температуре. Кроме того, часть аминокислот ответственна за подавление изомеризаций, отличных от all-E /13Z, например от all-E- к 7Z-, 9Z-, 11Z-ретиналю. Остальная часть полипептидной цепи обеспечивает канал протонного транспорта или экранирует фотохромную внутреннюю группу от влияний внешней среды. Взаимная топография образованных полипептидной цепью БР элементов вторичной структуры после поглощения молекулой хромофора кванта света изменяется, в результате чего формируется канал трансмембранного переноса протонов из цитоплазмы во внешнюю среду. Однако молекулярный механизм светозависимого транспорта до сих пор неизвестен.
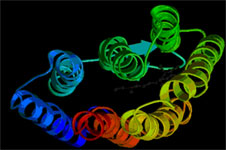 Рис.2.
Рис.2.
Схематическая модель трехмерной (пространственной) структуры БР (PDB Index 1FBB).Семь a-спиралей формируют хромофорную полость и трансмембранный канал переноса протона.
БР содержится в клеточной мембране H. salinarum - галофильной архебактерии, которая живет и размножается в соленых болотах и озерах, где концентрация NaCl может превышать 4 М, что в 6 раз выше, чем в морской воде (~ 0,6 М). Этот уникальный белок во многом подобен зрительному белку родопсину, хотя их физиологические функции различны. В то время как зрительный родопсин действует как первичный фоторецептор, который обеспечивает темновое зрение большинства позвоночных животных, физиологическая роль БР заключается в том, чтобы давать возможность галобактериям действовать как факультативным анаэробам в случае, когда парциальное давление кислорода в окружающей среде мало. Белок функционирует как светозависимый протонный насос, который обеспечивает образование электрохимического градиента протонов на поверхности мембраны клетки, который, в свою очередь, служит для аккумулирования энергии. Первичная работа, производимая градиентом, заключается в синтезе АТФ через анаэробное (фотосинтетическое) фосфорилирование и, в этом случае, представляет собой классический пример хемиосмотической гипотезы Митчелла об окислительном фосфорилировании. Когда освещение отсутствует, а парциальное давление кислорода высоко, бактерии возвращаются к аэробному окислительному фосфорилированию (Lanyi J.K., 1978). Клетки H. salinarum содержат также два так называемых сенсорных родопсина (СР I и СР II), которые обеспечивают положительный и отрицательный фототаксис. Различные длины волн считываются СР I и СР II как детекторными молекулами, что вызывает каскад сигналов, управляющих жгутиковым двигателем бактерии. При помощи такого элементарного процесса светового восприятия микроорганизмы самостоятельно перемещаются в свет подходящего спектрального состава. Кроме того, в клетках имеется галородопсин (ГР), представляющий собой светозависимый насос ионов Cl–. Его основная функция - транспорт в клетку ионов хлора, которые постоянно теряются бактерией, перемещаясь в направлении изнутри —> наружу под действием электрического поля, создаваемого БР. Механизм действия ГР неясен. Предполагается, что Cl– связывается с положительно заряженным четвертичным азотом протонированного Шиффова основания, а изомеризация ретиналя из all-E в 13Z-форму вызывает перемещение этого азота с прикрепленным к нему ионом Cl– от входного к выходному Cl– – проводящему пути.
Дополнительно
Технология выращивания сахарной свеклы в Сумской области
Сахарная свекла - важная техническая культура, корнеплод которой
достигает 500г и больше, содержит 19-22% сахара и более, является основным
сырьем для сахарной промышленности. Кроме сахара, в процессе переработки
корнеплодов получают ценные дополнительные продукты - мелясу и жом. Ботва
сахарной св ...
Расчет релаксационного генератора на ИОУ
Разработать и рассчитать
релаксационный генератор на ИОУ
(интегральной схеме операционного
усилителя) в соответствии с данными, представленными:
·
вид генератора - мультивибратор
·
режим работы – автоколебательный
·
период следования импульсов Т, мс – 0.09
· ...