Маслянокислое брожение
|
|
|
Рис. 57. Пути превращения пирувата в маслянокислом брожении, осуществляемом Clostridium butyricum: Ф1 — пируват:ферредоксиноксидоредуктаза; Ф2 — ацетил-КоА-трансфераза (тиолаза); Ф3 — (3-оксибутирил-КоА-дегидрогеназа; Ф4 — кротоназа; Ф5 — бутирил-КоА-дегидрогеназа; Ф6 — КоА-трансфераза; Ф7 — фосфотрансацетилаза; Ф8 — ацетаткиназа; Ф9 — гидрогеназа; Фдок — окисленный; Фд-H2 — восстановленный ферредоксин; ФН — неорганический фосфат |
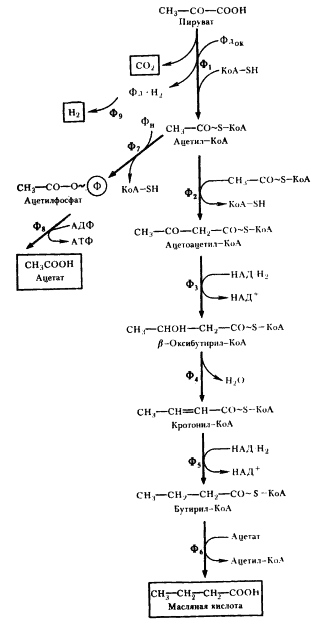
Следующий вариант решения донор-акцепторной проблемы на базе гликолитически образованного пирувата представляет собой маслянокислое брожение. Новое в маслянокислом брожении — возникновение реакций конденсации типа С2 + С2 ® С4, в результате чего образуется С4-акцепторная кислота. Судьба этой кислоты различна и определяется необходимостью акцептирования водорода с НАД-H2, освобождающегося в процессе брожения, а это в свою очередь тесно связано с оттоком водорода на конструктивные процессы. В качестве конечных C4-продуктов в процессе брожения возникают соединения различной степени восстановленности. Характерным C4-продуктом брожения является масляная кислота. Осуществляют такой тип брожения многие бактерии, относящиеся к роду Clostridium.
Типичными представителями клостридиев, осуществляющих маслянокислое брожение, являются C. butyricum и C. pasteurianum. Они сбраживают сахара с образованием масляной и уксусной кислот, CO2 и H2 (рис. 57). Превращение глюкозы до пирувата осуществляется по гликолитическому пути. Следующая реакция — разложение пирувата до ацетил-КоА и CO2, сопровождающееся образованием восстановленного ферредоксина (Фд). Реакция катализируется ферментом пируват: ферредоксиноксидоредуктазой и является ключевой в маслянокислом брожении. Особенности реакции — участие в ней белков, содержащих негемовое железо и кислотолабильную серу (FeS-белки)45.
К FeS-белкам относится группа белков, участвующих в процессах электронного транспорта (ферредоксины), и ряд ферментов, катализирующих окислительно-восстановительные реакции. Установлено, что FeS-белки являются ключевыми в таких важных клеточных процессах, как фотосинтез, дыхание, азотфиксация, фиксация CO2.
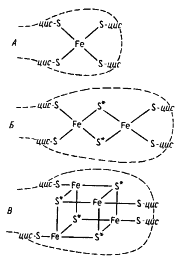
Отличительная особенность FeS-белков — строение их активного центра, содержащего негемовое железо, связанное нековалентными связями с кислотолабильной серой и серой, входящей в состав цистеиновых остатков пептидной цепи. Разные типы железосероцентров (FeS-центры) широко распространены в клетках. Простейший из них содержит один атом железа, нековалентно связанного в молекуле белка, получившего название рубредоксина, с четырьмя остатками цистеина (рис. 58, А)
|
|
|
Рис. 58. Железосероцентры FeS-белков. Железосероцентр рубредоксина (А); предполагаемые модели железосероцентров Fe2S2-типа (Б) и Fe4S4-типа (В). Звездочкой отмечена неорганическая кислотолабильная сера; прерывистой линией обозначена полипептидная цепь; цис — цистеин |
Обнаруженный у C. pasteurianum рубредоксин имеет окислительно-восстановительный потенциал около — 57 мВ и участвует в реакциях одноэлектронного переноса, в основе которого лежит переход железа:
Fe2+ « Fe3+.
Остальные FeS-белки имеют более сложно организованные FeS-центры, в состав которых входит также неорганическая кислотолабильная сера. Известны Fe2S2-центры (содержат по два атома железа и неорганической серы), Fe3S3- и Fe4S4-центры (рис. 58, Б, В). FeS-белки могут содержать один или более центров в молекуле. У большинства FeS-содержащих ферментов помимо FeS-центров в молекуле имеются и иные кофакторы: металлы (молибден, селен), хромофорные группы (флавин, гемы, птеридины), витамины (табл. 16).
Клостридии содержат ферредоксины с 1 — 2 центрами Fe4S4-типа и молекулярной массой 6000 — 7000 Да.
В зависимости от особенностей строения FeS-центров ферредоксины могут осуществлять одновременный перенос одного или двух электронов. Окислительно-восстановительный потенциал ферредоксинов находится в диапазоне от — 490 до — 310 мВ, однако описаны FeS-белки, окислительно-восстановительный потенциал которых высоко положителен (около + 350 мВ).
Ферредоксины играют центральную роль в метаболизме клостридиев, сопрягая катаболические процессы с биосинтетическими реакциями (рис. 59).
Таблица 16. Железосеросодержащие ферменты эубактерий
|
Простетическая группа |
Ферменты |
|
FeS-центры |
гидрогеназа и др. |
|
FeS-центры + тиаминпирофосфат |
пируват:ферредоксин-оксидоредуктаза |
|
FeS-центры + флавин |
сукцинатдегидрогеназа, НАД(Ф)-H2-дегидрогеназа, глутаматсинтетаза и др. |
|
FeS-центры + гем |
диссимиляционная сульфитредуктаза |
|
FeS-центры + молибден |
нитрогеназа, диссимиляционная нитратредуктаза, формиатдегидрогеназа и др. |
|
FeS-центры + два и более дополнительных кофактора |
ассимиляционная сульфитредуктаза, ксантиндегидрогеназа и др. |
Дополнительно
Распространение дифиллоботриоза в Костромской области и борьба с ним
Дифиллоботриоз плотоядных -
антропозооноз, имеющий очаговое распространение, вызывается различными видами
лентецов из рода Diphyllobothrium, среди которых
наиболее распространен лентец широкий -
Diphyllobothrium latum. Болеют собака, кошка, лисица, песец, куница, а также
человек.
Болезнь часто ...
Австрийская школа и теория предельной полезности
“Австрийская школа” возникла в 70-х годах 19-в.,
которые характеризовались дальнейшим ростом капитализма и обострением его
противоречий. На основе растущей концентрации производства в 70-х годах начали
возникать первые кап. монополии. Австрийская школа оспаривала учение Маркса, и
в авангарде этог ...