Маслянокислое брожение
|
|
|
Рис. 59. Роль ферредоксина в метаболизме клостридиев: 1 — пируват:ферредоксин-оксидоредуктаза; 2 — гидрогеназа; 3 — ферредоксин: НАД (Ф)-оксидоредуктаза; 4 — формиатдегидрогеназа; 5 — ксантиндегидрогеназа; 6 — нитрогеназа; 7 — ферредоксин:CO2-оксидоредуктаза (возможно, это формиатдегидрогеназа, катализирующая реакцию 4 в обратном направлении); 8 — реакция 3, протекающая в обратном направлении; 9 — сульфатредуктаза; 10 — реакция 1, протекающая в обратном направлении; 11 — нитратредуктаза |
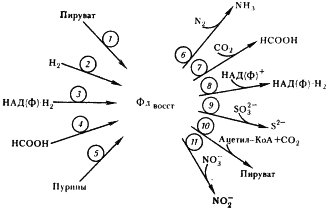
Объясняется это тем, что у клостридиев (как и других облигатных анаэробов) физиологические реакции в клетке всегда протекают при отрицательных окислительно-восстановительных потенциалах. В этих условиях FeS-белки, имеющие общий отрицательный окислительно-восстановительный потенциал, особенно пригодны для функционирования в составе ферментов и в качестве переносчиков электронов.
Образующийся в реакции восстановленный ферредоксин поставляет электроны для восстановления N2, протонов (H+), CO2 и НАДФ+, а последующее превращение ацетил-КоА приводит к синтезу АТФ в реакции субстратного фосфорилирования.
Путь, ведущий к синтезу масляной кислоты, начинается с реакции конденсации двух молекул ацетил-КоА (см. рис. 57). Образовавшийся ацетоацетил-КоА восстанавливается в b-оксибутирил-КоА. Источником электронов в этой реакции и дальше на пути синтеза масляной кислоты служат молекулы НАДxH2, образующиеся при окислении 3-ФГА в 1,3-ФГК (см. рис. 53).
Дальнейшее превращение заключается в отщеплении от молекулы (3-оксибутирил-КоА молекулы воды, что приводит к образованию соединения с двойной углеродной связью. Кротонил-КоА ферментативно восстанавливается в бутирил-КоА. Масляная кислота образуется в реакции переноса кофермента А с молекулы бутирил-КоА на ацетат. Эта реакция более "выгодна" для клетки, так как не приводит к потере энергии (в отличие от реакции простого гидролиза). Образующийся в реакции ацетил-КоА возвращается в метаболический поток и может быть использован для синтеза АТФ (реакция 7 на рис. 57) или же вновь участвовать в последовательности реакций, ведущих к синтезу масляной кислоты (реакции 2 — 6, там же).
Разобранный выше путь, завершающийся синтезом масляной кислоты, не связан с получением клеткой энергии, поскольку ни на одном из этапов не происходит образования АТФ. Единственное назначение метаболических превращений ацетил-КоА по этому пути — акцептирование электронов, переносимых на НАД+ в процессе гликолитического метаболизирования глюкозы: две молекулы НАД-H2 образуются на этапе гликолиза, и на двух этапах превращений ацетил-КоА до масляной кислоты происходит потребление водорода с НАД-H2.
В связи с этим особо важное значение приобретает превращение ацетил-КоА, ведущее к синтезу ацетата, поскольку именно с этим путем связано дополнительное получение клостридиями энергии в процессе маслянокислого брожения. Процесс включает несколько ферментативных реакций (см. рис. 57). Сначала имеет место окислительное декарбоксилирование пировиноградной кислоты, катализируемое пируват:ферредоксиноксидоредуктазой. Далее с помощью гидрогеназы происходит выделение молекулярного водорода с восстановленного ферредоксина.
Гидрогеназы — одна из групп FeS-содержащих ферментов, катализирующих реакции поглощения и выделения молекулярного водорода, обнаружены у разных групп эубактерий: облигатных анаэробов и аэробов, факультативных форм, у хемо- и фототрофных организмов. Различаются строением молекулы, природой доноров и акцепторов электронов, с которыми взаимодействуют, локализацией в клетке, выполняемыми функциями. Но все гидрогеназы катализируют реакцию H2 ® 2H+ + 2e– .
Гидрогеназа C. pasteurianum, один из наиболее детально изученных ферментов, — белок с молекулярной массой примерно 60 000 Да, представленный одной субъединицей. В молекуле содержатся три центра типа Fe4S4. Донором (акцептором) электронов клостридиальной гидрогеназы служит ферредоксин.
При разрушении клеток C. pasteurianum гидрогеназная активность проявляется только в растворимой фракции: в периплазматическом пространстве и цитоплазме. Гидрогеназа, локализованная в периплазматическом пространстве, катализирует необратимую реакцию поглощения H2. Находящаяся в цитоплазме гидрогеназа способна катализировать реакции как поглощения, так и выделения H2. У клостридиев она входит в состав ферментного комплекса, осуществляющего окислительное декарбоксилирование пирувата (см. рис. 57).
Основная функция гидрогеназ клостридиев (и других облигатных анаэробов) заключается в избавлении от избытка образующихся в катаболических реакциях восстановительных эквивалентов (электронов), которые переносятся на H+ и удаляются из клетки в виде молекулярного водорода.
Дополнительно
Биологическое время и его моделирование в квазихимическом пространстве
Методология построения теории времени естественных объектов, детально
изложена [1, 2]. В данной работе рассмотрены компоненты этой теории на примере
клеточной популяции.
...
Галактика как уровень мегамира
Актуальность,
цели и задачи ответа по настоящей контрольной работе будут обусловлены
следующими положениями. Нас интересует не только звездное население того дома,
в котором мы живем. Нас интересует и архитектура этого дома и его размеры;
интересует, как его обитатели расселены, где жилищная тесно ...