Окислительный пентозофосфатный путь
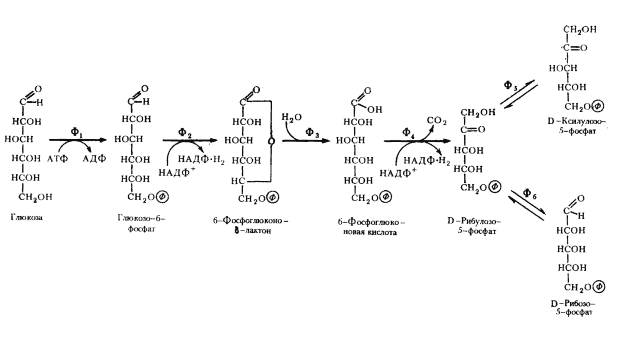
Схема начальных этапов окислительного пентозофосфатного пути представлена на рис. 64. Первая реакция заключается в фосфорилировании глюкозы с помощью АТФ и превращении ее в метаболически активную форму глюкозо-6-фосфата, аналогично тому, что имеет место на первом этапе гликолиза. Следующий этап заключается в дегидрировании глюкозо-6-фосфата, катализируемом глюкозо-6-фосфатдегидрогеназой. Особенность реакции в том, что в ней участвует НАДФ+ в качестве акцептора водорода. Образовавшийся продукт реакции очень нестоек и спонтанно или с помощью фермента лактоназы гидролизуется с образованием 6-фосфоглюконовой кислоты, которая подвергается окислительному декарбоксилированию, катализируемому фосфоглюконатдегидрогеназой. Эта реакция приводит к образованию соответствующего пентозофосфата, НАДФ-H2 и выделению CO2. Рибулозо-5-фосфат обратимо превращается в ксилулозо-5-фосфат и рибозо-5-фосфат с участием ферментов фосфопентозоэпимеразы и фосфопентозоизомеразы соответственно.
Суммарно весь процесс можно представить в виде следующего уравнения:
глюкозо-6-фосфат + 2НАДФ+ ® рибозо-5-фосфат + CO2 + 2НАДФ-Н2.
Как видно, на этом этапе образуются 2 молекулы НАДФ-H2, которые могут потребляться в восстановительных биосинтетических процессах, и молекула рибозо-5-фосфата, используемого в синтезе нуклеиновых кислот и пентозосодержащих коферментов49. Примечательно, что ни на одном из окислительных этапов не синтезируется АТФ.
Первоначально окислительный пентозофосфатный путь возник, вероятно, для обеспечения эубактерий пентозами. В этом случае возникновение только трех новых ферментов (глюкозо-6-фосфатдегидрогеназы, лактоназы и фосфоглюконатдегидрогеназы) уже приводило к синтезу пентоз. Поскольку к этому времени функционировали изомеразные ферменты гликолитического пути (см. рис. 53), формирование фосфопентозоизомеразы произошло довольно легко. Действительно, при определенных условиях окислительный пентозофосфатный путь на этом завершается.
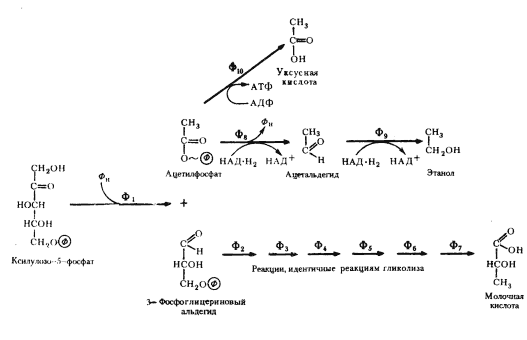
Дальнейшее его развитие, вероятно, связано с энергетическими потребностями клетки. Меньшей части образующегося рибозо-5-фосфата оказалось достаточно для удовлетворения всех потребностей клетки в пентозах. Остальная часть синтезируемого пентозофосфата была субстратом, хранившим в себе большие запасы энергии. Способность использовать в энергетических целях этот субстрат связана с возникновением двух ферментов: фосфопентозоэпимеразы, катализирующей превращение рибулозо-5-фосфата в ксилулозо-5-фосфат (рис. 64), и пентозофосфокетолазы, катализирующей расщепление ксилулозо-5-фосфата на 3-ФГА и ацетилфосфат (рис. 65).
Использование в качестве источника энергии в анаэробных условиях пентозных субстратов, образуемых в окислительном пентозофосфатном пути, свойственно группе гетероферментативных молочнокислых бактерий, для которых характерно образование в качестве конечных продуктов брожения ряда органических соединений: молочной и уксусной кислот, этилового спирта, глицерина, CO2 и др. Этим гетероферментативные молочнокислые бактерии отличаются от гомоферментативных, почти полностью сбраживающих гексозы по гликолитическому пути в молочную кислоту.
|
|
|
Рис. 64. Окислительный пентозофосфатный путь (начальные этапы): Ф1 — гексокиназа; Ф2 — глюкозо-6-фосфатдегидрогеназа; Ф3 — лактоназа; Ф4 — фосфоглюконатдегидрогеназа (декарбоксилирующая); Ф5 — фосфопентозоэпимераза; Ф6 — фосфопентозоизомераза (по Dagley, Nicholson, 1973) |
Изучение механизмов образования конечных продуктов брожения гетероферментативными молочнокислыми бактериями обнаружило, что они связаны с дальнейшими различными путями метаболизирования C2- и C3-фрагментов фосфокетолазной реакции. 3-ФГА претерпевает ряд ферментативных превращений, идентичных таковым гликолитического пути, и через пируват превращается в молочную кислоту. Судьба двухуглеродного фрагмента различна: двухступенчатое восстановление ацетилфосфата приводит к накоплению в среде этанола; окислительный путь превращения ацетилфосфата завершается образованием уксусной кислоты (рис. 65).
|
|
|
Рис. 65. Гетероферментативное молочнокислое брожение: Ф1 — пентозофосфокетолаза; Ф2 — 3-ФГА-дегидрогеназа; Ф3 — фосфоглицераткиназа; Ф4 — фосфоглицеромутаза; Ф5 — енолаза; Ф6 — пируваткиназа; Ф7 — лактатдегидрогеназа; Ф8 — ацетальдегиддегидрогеназа; Ф9 — алкогольдегидрогеназа; Ф10 — ацетаткиназа (по Schlegel, 1972) |
Дополнительно
Счетчики ядерного излучения
Реальная
перспектива использования человеком огромных энергий, скрытых в недрах атома,
появилась впервые в 1939 году. На сегодняшний день широкое практическое применение
получают различного рода ядерные излучения, несмотря на то, что они опасны для
организма человека и в то же время неощущаемы, п ...
Современная судовая газотурбинная установка
Современная
судовая газотурбинная установка (ГТУ) успешно конкурирует с аналогичными по
назначению паротурбинными и дизельными. От последних она выгодно отличается
компактностью и малой удельной массой, маневренностью и высокой
ремонтопригодностью, лучшей приспособленностью к автоматизации ...