Окислительный пентозофосфатный путь
Преобладание в ферментационной среде того или иного продукта зависит от вида культуры, условий культивирования и фазы развития. Гетероферментативные молочнокислые бактерии Leuconostoc mesenteroides сбраживают глюкозу в молочную кислоту, этанол и CO2 по следующему уравнению:
C6H12O6 ® CH3-CHOH-COOH + CH3-CH3-CH2OH + CO2
У других гетероферментативных молочнокислых бактерий больший удельный вес занимают процессы, ведущие к накоплению уксусной кислоты. Образование уксусной кислоты из ацетилфосфата сопряжено с синтезом АТФ. Если брожение идет с образованием этанола, то выход энергии равен 1 молекуле АТФ на молекулу сброженной глюкозы; если образуется уксусная кислота, то общий энергетический баланс процесса составляет 2 молекулы АТФ на молекулу глюкозы, т. е. такой же, как при гликолизе.
Окислительный пентозофосфатный путь функционирует в качестве единственного пути сбраживания углеводов у облигатных гетероферментативных молочнокислых бактерий. Эти бактерии лишены ключевых ферментов гликолитического пути, например альдолазы и триозофосфатизомеразы. Большинство молочнокислых бактерий имеют два пути сбраживания углеводов: гликолитический и окислительный пентозофосфатный. Сбраживание гексоз, как правило, протекает по гликолитическому пути, а пентоз — по окислительному пентозофосфатному. Это имеет место, например у Lactobacillus plantarum. Ферменты окислительного пентозофосфатного пути обнаружены у клостридиев.
Таким образом, возникнув сначала как механизм синтеза клеткой C5-соединений, т. е. для выполнения узкой специфической задачи, этот путь получил дальнейшее развитие и стал выполнять дополнительную функцию снабжения эубактерий энергией в анаэробных условиях. Субстратная база для окислительного пентозофосфатного пути позднее была расширена, так как он стал использоваться и для сбраживания пентоз биогенного происхождения, накапливавшихся в окружающей среде.
Но на этом пути эволюционное развитие окислительного пентозофосфатного пути расщепления углеводов не остановилось. Была сформирована последовательность реакций, "замыкающая" этот путь в цикл, в результате чего стала возможной полная деградация молекулы сахара. Исходными субстратами на этом пути служат пентозы, образующиеся из рибулозо-5-фосфата, ксилулозо-5-фосфата и рибозо-5-фосфата (см. рис. 64). При участии двух дополнительных ферментов — транскетолазы и трансальдолазы — осуществляется перенос C2- и C3-фрагментов между изомерными пентозо-5-фосфатами и продуктами их взаимопревращений (рис. 66). Сначала транскетолаза переносит C3-фрагмент от молекулы ксилулозо-5-фосфата на молекулу рибозо-5-фосфата, в результате чего образуется C7-сахар и C3-сахар — 3-ФГА. 3-ФГА, образующийся в транскетолазной реакции и, как известно, представляющий собой промежуточный продукт гликолитического пути, является первой точкой, в которой пересекаются эти пути.
Далее трансальдолаза действует на продукты транскетолазной реакции, перенося C3-фрагмент от молекулы C7-сахара на C3-молекулу — 3-ФГА. В результате образуются молекулы C6-и C4-сахара. Один из продуктов реакции — фруктозо-6-фосфат является промежуточным соединением гликолитического пути, поэтому данная реакция есть вторая точка пересечения обоих путей углеводного метаболизма. Наконец, транскетолаза осуществляет перенос C2-фрагмента от молекулы D-ксилулозо-5-фосфата на молекулу C4-сахара по той же схеме, что и в первой транскетолазной реакции.
|
|
|
Рис. 66. Окислительный пентозофосфатный путь (конечные этапы): Ф1 — транскетолаза; Ф2 — трансальдолаза; кружками обведены C2 — гликольальдегидная и C3 — диоксиацетоновая группы; в квадраты заключены C3 — 3-фосфоглицериновый альдегид (3-ФГА), C4 — D-эритрозо-4-фосфат, C6 — D-фруктозо-6-фосфат, C7 — D-седогептулозо-7-фосфат (по Schlegel, 1972) |
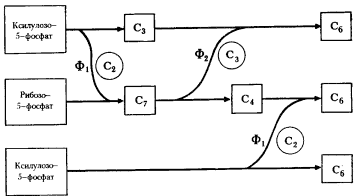
Итог этих взаимопревращений таков: из 3 молекул пентозофосфата синтезируются 2 молекулы фруктозо-6-фосфата и 1 молекула 3-ФГА. Фруктозо-6-фосфат ферментативно превращается в глюкозу, и 2 молекулы глюкозы снова возвращаются в цикл. 2 молекулы 3-ФГА также могут конденсироваться с образованием 1 молекулы глюкозы. В результате функционировавания полного окислительного пентозофосфатного пути из 6 поступающих в него молекул глюкозы 5 молекул ревосстанавливаются, а одна полностью окисляется до CO2, что приводит к восстановлению 12 молекул НАДФ+ до НАДФ-H2. Это можно представить в виде следующего уравнения:
6 глюкозо-6-фосфат + 12НАДФ+ ® 5 глюкозо-6-фосфат + 6CO2 + 12НАДФ-H2 + ФН.
Таким образом, окислительный пентозофосфатный путь может служить циклическим механизмом полной деградации углеводов, при этом водород, отщепленный от глюкозы, поступает в электронтранспортную цепь и переносится на O2.
Дополнительно
Современная судовая газотурбинная установка
Современная
судовая газотурбинная установка (ГТУ) успешно конкурирует с аналогичными по
назначению паротурбинными и дизельными. От последних она выгодно отличается
компактностью и малой удельной массой, маневренностью и высокой
ремонтопригодностью, лучшей приспособленностью к автоматизации ...
Методы оценки близости допредельных и предельных распределений статистик
Рассматривается проблема оценки близости предельных
распределений статистик и распределений, соответствующих конечным объемам
выборок. При каких объемах выборок уже можно пользоваться предельными
распределениями? Каков точный смысл термина "можно" в предыдущей фразе?
Основное внимание уд ...