РНК, ДНК, белки и мембраны.
Хорошими кандидатами на роль энергетического резерва первичных клеток могли быть неорганические пиро- и полифосфаты. Они и сегодня играют эту роль у некоторых видов живых существ. Например, в клетках грибов полифосфаты образуются из АТФ в условиях избытка энергетических ресурсов и расщепляются, давая АТФ, при дефиците источников энергии. Однако в подавляющем большинстве дошедших до нас организмов функцию легко мобилизуемого энергетического резерва выполняют не полифосфаты, а углеводы. Их синтез за счет энергии АТФ (гликогенез) представляет собой длинную последовательность реакций, намного более сложную, чем синтез полифосфатов из АТФ.
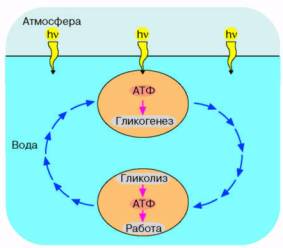
Рис. 2. Энергетика первичной живой клетки, основанная на "адениновом" фотосинтезе. Предполагается, что ультрафиолетовые кванты, достигая поверхности первичного океана, использовались клеткой для синтеза АТФ, который запускал синтез углеводородов (гликогенез). Клетка, увлекаемая потоком океанской воды с поверхности на некоторую глубину, оказывалась вне досягаемости ультрафиолетового облучения. Здесь происходило расщепление накопленных углеводов и синтез АТФ, использовавшегося для совершения клеткой различных типов полезной работы.
Преимущество углеводов перед полифосфатами состоит в том, что в них запасены не только энергия, но и "строительный материал". Расщепление углеводов (гликолиз) дает помимо АТФ карбоновые кислоты, такие, как пировиноградная кислота, которая может использоваться клеткой при биосинтезе самых разнообразных соединений.
Описаны два основных типа гликолиза. В одном случае (спиртовое брожение) конечными продуктами расщепления углеводов оказываются этиловый спирт и углекислый газ — вещества, легко проникающие через мембрану клетки. Это обстоятельство имеет как преимущества (нет проблемы переполнения клетки конечными продуктами гликолиза), так и недостатки (трудно вернуться назад, к углеводу, если конечные продукты уже вышли из клетки и разбавились в океане внешней среды).
Указанный недостаток отсутствует во втором, сегодня гораздо более распространенном типе гликолиза, когда конечным продуктом оказывается молочная или какая-либо другая карбоновая кислота. Молочная кислота не проникает через мембрану, не покидает пределы клетки и потому может быть использована клеткой для ресинтеза углеводов, когда возникает такая возможность. Неудачно лишь то, что молекулы молочной кислоты, образуясь, диссоциируют с образованием ионов лактата и водорода. Последние также не могут пройти через мембрану, остаются в клетке и закисляют ее содержимое, Закисление, если его не предотвратить, должно привести к гибели клетки из-за кислотной денатурации белков. Решение этой проблемы описано в следующем разделе.
Дополнительно
Внутренняя структура протона и новый способ получения энергии
Протон был открыт в начале 20-х г.г. в экспериментах с альфа-частицами. В
опытах по рассеянию на протонах электронов и гамма-квантов были получены
достоверные доказательства существования некой внутренней структуры у этой
частицы. В 1970 г. в Стенфордском центре линейного ускорителя (СЛАК) удалось ...
Детские дошкольные учреждения – сады-ясли
Двадцатое столетие для
рядя стран Европы характерно процессами интенсивной урбанизации в связи с
индустриализацией производства и соответствующим размахом градостроительной
деятельности.
В нашей стране процесс
урбанизации привел к исключительно острой проблеме обеспечения жилищем и
общественны ...