Возникновение фотосинтеза, использующего видимый свет.
Устройство бактериородопсина намного проще
системы хлорофилльного фотосинтеза. Белковая часть бактериородопсина представляет собой одну полипептидную цепь средней длины, которая не содержит других коферментов и простетических групп, кроме ретинштя. Бактериородопсин чрезвычайно устойчив: без потери активности его можно кипятить в автоклаве при + 130°С, изменять содержание NaCl в омывающем мембрану растворе от нуля до насыщения, в широких пределах менять рН этого раствора. Более того, можно удалить выступающие из мембраны концевые участки полипептид-ной цепи и даже расщепить эту цепь в одном месте по середине без ущерба для активности насоса. В то же время эффективность бактериородопсина как преобразователя энергии сравнительно низка: всего 20% энергии светового кванта превращается в ΔμΗ+. При этом на один поглощенный квант через мембрану переносится один ион Н+.
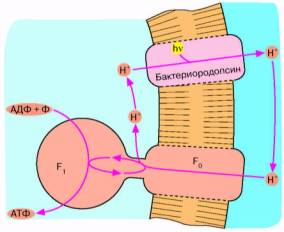
Рис. 4. Бактериородопсиновый фотосинтез со-
лелюбивых архебактерий. Ионы Н* откачиваются из клетки бактериородопсином - белком, содержащим ретиналь в качестве хромофора, то есть группировки, поглощающей видимый свет. Ионы Н* возвращаются в клетку, двигаясь "под гору" через Н*-АТФазный комплекс F0F,. При этом оказывается, что 1-Г-АТФаза катализирует обратную реакцию, то есть синтез АТФ, а не его гидролиз
Хлорофилльный фотосинтез
Хлорофилльный фотосинтез отличается от бак-териородопсинового большей эффективностью использования светового кванта. Он устроен таким образом, что либо на каждый квант переносится через мембрану не один, а два иона Н+, либо помимо транспорта Н+ происходит запасание энергии в форме углеводов, синтезируемых из СО2 и Н2О. Вот почему бактериородопсиновый фотосинтез был оттеснен эволюцией с авансцены. Он сохранился только у бактерий, живущих в экстремальных условиях, где более сложный и менее устойчивый Хлорофилльный фотосинтез, по-видимому, просто не может существовать.
Хлорофилльный фотосинтез катачизируется ферментной системой, включающей несколько белков. Квант света поглощается хлорофиллом, молекула которого, перейдя в возбужденное состояние, передает один из своих электронов в фотосинтетическую цепь переноса электронов. Эта цепь представляет собой последовательность окислительно-восстановительных ферментов и коферментов, находящихся во внутренней мембране бактерий или хлоропластов растений, где локализованы также белки, связанные с хлорофиллом. Компоненты цепи содержат, как правило, ионы металлов с переменной валентностью (железо, медь, реже марганец или никель). При этом железо может входить в состав тема (в таком случае белки называются цито-хромами). Большую роль играют также негемовые железопротеиды, где ион железа связан с белком через серу цистеина или реже азот гистидина. Помимо ионов металлов роль переносчиков электронов играют производные хинонов, такие, как убихинон, нластохинон и витамины группы К.
Перенос по цепи электрона, отнятого от возбужденного хлорофилла, завершается по-разному в зависимости от типа фотосинтеза. У зеленых бактерий, использующих комплекс хлорофилла и белка, называемый фотосистемой 1 (рис. 5, а), продуктом оказывается НАДН, то есть восстановленная форма НАД+. Восстанавливаясь, то есть присоединяя два электрона, НДЦ+ связывает также один Н+. В дальнейшем образованный таким образом НАДН окисляется, передавая свой водород на различные субстраты биосинтезов.
Что касается хлорофилла, окисленного цепью, то у зеленых серных бактерий он получит недостающий электрон от сероводорода (H2S). В результате образуются также элементарная сера и ион Н+. Белок, окисляющий H2S, расположен на внешней поверхности бактериальной мембраны, а белок, восстанавливающий НАД+, — на внутренней ее поверхности. Вот почему оказывается, что запускаемый светом перенос электронов от H,S к НАД+ образует ионы Н+ снаружи и потребляет их внутри бактерии. При этом внутренний объем клетки заряжается отрицательно относительно внешнего. Тем самым создается ДДН+, которая потребляется Н+-АТФ-синтазой (комплексом факторов F0 и FJ, образующей АТФ при переносе ионов Н "под гору", то есть снаружи внутрь.
Дополнительно
Технология выращивания сахарной свеклы в Сумской области
Сахарная свекла - важная техническая культура, корнеплод которой
достигает 500г и больше, содержит 19-22% сахара и более, является основным
сырьем для сахарной промышленности. Кроме сахара, в процессе переработки
корнеплодов получают ценные дополнительные продукты - мелясу и жом. Ботва
сахарной св ...
Детские дошкольные учреждения – сады-ясли
Двадцатое столетие для
рядя стран Европы характерно процессами интенсивной урбанизации в связи с
индустриализацией производства и соответствующим размахом градостроительной
деятельности.
В нашей стране процесс
урбанизации привел к исключительно острой проблеме обеспечения жилищем и
общественны ...