Методология.
Существуют два различных, но, как станет очевидным, взаимозависимых пути, по которым развивалась адаптационистская программа (5).
Метод 1 (апостериорный!сравнительный подход). При этом методе изучают и сравнивают признаки у близких видов в разные периоды истории данной группы (то есть по ископаемым остаткам) или же — в современной фауне — у видов, занимающих различные экологические ниши. Затем (после такого изучения) дается объяснение обнаруженных различий в адаптивном плане. В отношении различий, коррелированных со временем, принимается допущение, что изменения в признаках следуют за изменениями экологических условий и (или) представляют собой прогрессивные адаптивные усовершенствования. Так, например, направления в эволюции лошадей (рис. 3.1) объясняют переходом их от жизни в лесах к жизни на равнинах. В процессе эволюции наблюдается: 1) увеличение общих размеров, в частности длины конечностей, которые при этом становятся и более стройными; 2) уменьшение числа пальцев до одного и разрастание «ногтя» этого оставшегося пальца с образованием копыта; 3) увеличение резцов в ширину и коренных зубов в высоту вместе с развитием сложных складок твердого цемента на их жевательной поверхности. Изменения 1 и 2 можно рассматривать как адаптации к быстрому передвижению по твердой почве, а изменение 3 — как адаптацию к питанию жесткой степной травой.
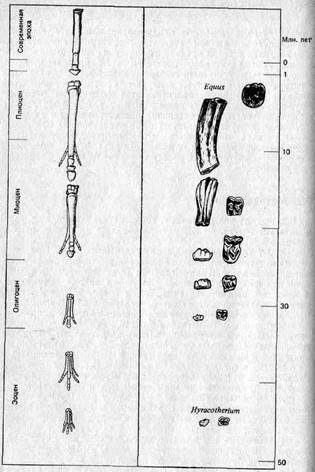
Рис. 3.1. Направления эволюции конечностей и коренных зубов у лошадей (объяснения см. в тексте).
Что касается изменений, коррелированных с пространством, то считается, что дивергенция признаков у популяций, принадлежащих к одному и тому же или к близким видам, произошла в результате различных давлений отбора, испытываемых этими популяциями в тех различных экологических условиях, в которых они находятся. Следовательно, нам важно иметь возможность точно определять различия в тех экологических условиях, под действием которых находятся организмы! Физиологическим экологам, изучающим прибрежную фауну, известно множество примеров близкородственных видов, обитающих в разных частях приливно-отливной зоны и поэтому в разной степени подверженных обсыханию в промежутке между двумя приливами и воздействию волн. Морфологические, физиологические и поведенческие различия между видами, обитающими в разных частях приливно-отливной зоны, во многих случаях можно объяснить как адаптации к этим экологическим различиям; один из таких примеров приведен в табл. 3.1. В этом случае большую устойчивость к высоким температурам и иссушающим условиям у Patella vulgata, обитающей в верхней части этой зоны, чем у P. aspera, обитающей в ее нижней части, можно рассматривать как приспособление к более частому и более длительному пребыванию в воздушной среде в промежутке между приливами. Кроме того, более низкую скорость потери воды в воздушной среде у P. vulgata по сравнению с P. aspera можно отнести за счет различий в форме раковины у этих двух видов (у P. vulgata поверхность, обсыхающая в воздушной среде, меньше); таким образом, морфологические различия поддаются объяснению с позиций адаптационизма.
Таблица 3.1. Некоторые аспекты физиологической экологии морских видов блюдечка (по данным Девиса (10,11))
|
Признак |
patella vulgata |
Patella aspera |
|
Зона обитания Выживаемость при t<30°C Скорость потери воды тканями Способность противостоять потере воды |
Верхняя Высокая Низкая Высокая |
Нижняя Низкая Высокая Низкая |
|
Форма раковины |
|
|
В связи с методом 1 возникает ряд проблем.
1. Не во всех случаях можно твердо установить, являются ли наблюдаемые различия генотипическими или лишь фенотипиче-скими. Так, например, различия в форме раковины, приведенные в табл. 3.1, можно отнести за счет прямого воздействия волн, то есть в этом случае, если бы оба вида развивались в одних и тех же условиях, никаких различий между ними не было бы. Подобного рода негенетические различия (называемые иногда фенотипической пластичностью) в данном контексте интереса не представляют. Отметим, однако, что фенотипическая пластичность может быть детерминирована генетически, а может быть адаптивной. Примеры исследований, в которых был проведен более точный генетический анализ, будут даны в разд. 3.3.
2. Наличие корреляции само по себе нельзя считать доказательством существования причинно-следственной зависимости. Возможно, что в возникновении различий участвуют какие-то скрытые переменные, играющие более важную роль. Возможна также, что различия в форме раковины у блюдечка обусловлены различиями в выеданий хищниками, а это последнее регулируется воздействием волн или обсыханием в промежутке между приливами. В таком случае различия в признаках следовало бы объяснять в свете того, в какой степени раковины определенной формы обеспечивают защиту от хищников, а не от обсыхания или воздействия волн.
Дополнительно
Становление детской журналистики и её влияние на психологию ребёнка
Русская пресса для юного читателя в
отличие от «взрослой» началась с журнала.
Детская журналистика в нашей стране
имеет богатую историю. До революции, в основном в Петербурге и Москве,
издавалось около трехсот детских и юношеских журналов. Одни из них выходили
десятилетиями, другие прекращали ...
Естественно-научные концепции развития микроэлектронных и лазерных технологий
Электроника - наука о взаимодействии электронов
с электромагнитными полями и о методах создания электронных приборов и
устройств (вакуумных, газоразрядных, полупроводниковых), используемых для
передачи, обработки и хранения информации. Возникла она в начале ХХ века. На ее
основе были созданы элект ...