От молекулы к клетке
Предпосылками для появления транскрипции у пробионтов было то, что в них уже производились молекулы АТФ и другие макроэрги, из которых синтезируются ДНК и РНК. Вполне могли появиться двухцепочечные фрагменты ДНК - будущий матричный материал. Но поскольку еще не было ферментов, требовались другие факторы, управляющие разделением двух цепей ДНК, синтезом РНК и восстановлением спаренной ДНК.
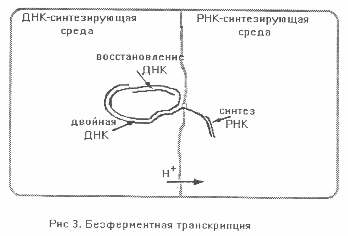
Мы предположили, что решающим фактором стало различие pH в соседних участках внутренней среды пробионта - так называемая компартментализация. Причиной параметрического различия сред был перенос протонов через каталитически активные пленки. (Аналогичные процессы в живых клетках - трансмембранные протонные переносы открыл английский ученый П.Митчел.) В одном из компартментов пробионта создавались условия, благоприятные для синтеза РНК, а в другом - ДНК. Транскрипция начиналась с рассоединения двойной спирали ДНК и переноса одной из ее цепей через мембрану, разделяющую две среды. Эта цепь становилась матрицей для синтеза РНК, а другая оставалась в прежнем компартменте и служила матрицей для восстановления парной ДНК (Рис. 3). Можно предположить, что и у живых клеток важным условием транскрипции служит компартментализация при переносе протонов через мембрану, содержащую рибосомы.
На третьем этапе абиогенеза благодаря матричному синтезу произошло наращивание генетического материала ДНК, количественное и качественное усовершенствование РНК. Возникла согласованность многих каталитических процессов и открылись новые перспективы усложнения пробионтов.
Так как УФ-энергия поступала только днем, ночью пробионты теряли активность и распадались, а каждое утро рождались заново. Это стало очередным эволюционным препятствием, для преодоления которого на последнем этапе абиогенеза нужны были принципиально новые “ночные” системы энергоснабжения. Как известно, у многих бактерий основой энергетики является гликолиз. Энергия, высвобождающаяся от частичного окисления некоторых органических соединений, направляется на фосфорилирование, т.е. на синтез АТФ и других макроэргов. Различные варианты подобного окислительного фосфорилирования служат важной частью энергоснабжения всех живых клеток.
Для перехода от ежедневных циклов ассимиляции и распада у пробионтов к круглосуточной жизни клетки требовался процесс, подобный гликолизу, направляющий химическую энергию органических молекул на синтез АТФ. Известно, что без ферментов он невозможен, т.к. доминирует обратный процесс - гидролиз АТФ. Ферменты образуются только из полипептидов, синтезированных матричным путем. Следовательно, зарождению первых клеток должен был предшествовать матричный синтез полипептидов.
В пробионтах уже существовали молекулярные системы безматричного синтеза полипептидов и матричного синтеза РНК. Имелись нуклеопротеидные комплексы, обладающие сродством с аминокислотами - предшественники тРНК. Оставалось преодолеть очередной эволюционный барьер - создать генетический код. Первоначально он мог быть значительно проще современного. Даже грубое кодирование основных типов аминокислот сказывалось на свойствах синтезированных макромолекул и, тем самым, давало пробионтам огромные селективные преимущества. Первые аналоги рибосом могли представлять собой относительно простые нуклеопротеидные комплексы, в которых контроль кодон-антикодонового соответствия иРНК и тРНК шел за счет переносов энергии при их контактах. Возможно, следы этих микропроцессов проявляются в работе рибосом всех живых клеток и могут быть выявлены экспериментально.
Попытки матричного синтеза полипептидов продолжались долгое время, не оказывая существенного влияния на эволюцию пробионтов. Но как только были синтезированы первые молекулы, способные связать какую-либо окислительную реакцию с фосфорилированием, матричные процессы приобрели огромную селективную ценность. Пробионты перешли в режим круглосуточной активности. Главным критерием отбора стало качество синтезируемых ферментов, их влияние на производство АТФ. Можно предположить, что первые ферменты катализировали окисление сахарного альдегида в фосфоглицериновую кислоту, сопряженное с переносом неорганического фосфата на АДФ. Позже по мере истощения запасов органики в первичном бульоне, цепочки ферментативного окислительного фосфорилирования удлинялись.
Дополнительно
Оборудование для механического обезвоживанья и сушки текстильных материалов
Сушка является самым распространенным технологическим
процессом красильно-отделочного производства. На многих отделочных фабриках
сушильное оборудование занимает приблизительно до 30 % производственных
площадей, потребляет до 40 % всего расходуемого тепла и до 30 % электроэнергии.
Одним из эффек ...
Детские дошкольные учреждения – сады-ясли
Двадцатое столетие для
рядя стран Европы характерно процессами интенсивной урбанизации в связи с
индустриализацией производства и соответствующим размахом градостроительной
деятельности.
В нашей стране процесс
урбанизации привел к исключительно острой проблеме обеспечения жилищем и
общественны ...