Неодарвинизм и популяционная генетика.
Рис. 2.8. Адаптивный ландшафт при различных частотах двух хромосомных инверсий у одного из представителей прямокрылых. BL — инверсия Blundell; TD —инверсия Tidbinbilla. (Shorrocks В., The Genesis of Diversity, Hodder a. Stoughton, London, 1978, по данным Lewontin R. C., White M. J. D., Evolution, 1960, 13, 561—564.)
На рис. 2.9 адаптивные ландшафты использованы для того, чтобы представить в качественном аспекте некоторые возможные эволюционные ситуации. (Следует помнить, что мы рассматриваем эти ландшафты сверху, то есть видим их контуры.) На рис. 2.9, А показано, чего следует ожидать в случае ослабления действия отбора или усиления мутационного процесса: область, занимаемая некой популяцией, распространяется по склонам ее адаптивного пика, и если это происходит достаточно интенсивно, то вид, к которому она принадлежит, будет вынужден занять еще один пик. На рис. 2.9, Б изображены результаты усиления отбора; в этом случае популяция оказывается сосредоточенной на высшем уровне своего адаптивного пика. Рис. 2.9, В иллюстрирует результаты изменений в среде; в этом случае рельеф ландшафта изменяется, так что адаптивный пик может «рухнуть» под тяжестью популяции, которая таким образом окажется в «долине». Затем под действием отбора популяция начнет подниматься на какой-либо близлежащий пик.
Как показывают рис. 2.9, Л и Б, для того чтобы популяция оставалась на занятом ею пике, необходим непрерывный отбор; такой отбор получил название стабилизирующего отбора: в результате, непрерывного отбора против мутантов популяция сохраняет стабильность в отношении как морфологических, так и физиологических признаков. Процесс В, хотя в его основе лежат те же механизмы, приводит к другим результатам: под его воздействием популяция приобретает новое адаптивное сочетание признаков. Такой отбор получил название направленного отбора.
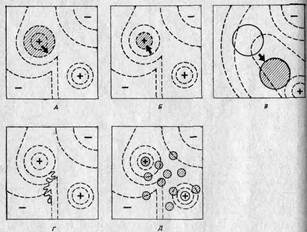
Рис. 2.9. Несколько адаптивных ландшафтов — вид сверху. А. Ослабление отбора или усиление мутационного процесса. Б. Стабилизирующий отбор. В. Направленный отбор. Г. Случайный дрейф-Д. Популяция, разделившаяся на демы. (Wadd-ington С. Н., An Introduction to Modern Genetics, George Alien a. Unvin Ltd, London, 1939.)
На рис. 2.9, Г представлена небольшая популяция. Здесь случайные события могут вызвать сдвиг в сторону от адаптивного пика, несмотря на отбор, и это та возможность, которую Сьюэлл Райт сформулировал вполне определенно. В принципе каждый индивидуум может случайно стать жертвой несчастного случая или же не найти себе брачного партнера, в результате чего он не внесет своего вклада в следующее поколение независимо от того, сколь хорошо он приспособлен. Поэтому гены, которые он несет, как бы хороши они ни были потенциально, не будут представлены в следующем поколении. Для большой популяции это не будет иметь существенного значения. Однако в небольших популяциях такие случайные события могут оказать серьезное воздействие на частоты генов, так как эти частоты определяются небольшим числом особей. При таких обстоятельствах популяция может медленно отойти от адаптивного пика; процессе этот получил название генетического дрейфа. Подобным же образом при резком сокращении численности популяции или же, что, в сущности, то же самое, при образовании несколькими особями-основателями новой популяции выборка генов из генофонда исходной популяции может произойти неслучайным образом. В результате будет иметь место некое начальное изменение, не связанное с отбором. Это явление известно под названием эффекта основателя.
На рис. 2.9, Д изображена большая популяция, разделенная на ряд субпопуляций, каждая из которых занимает части данного адаптивного ландшафта, несколько отличающиеся друг от друга. Такие популяции, вероятно, встречаются довольно часто. Степень отклонения каждой группы (в результате отбора или дрейфа) от среднего генетического состава популяции в целом зависит от интенсивности обмена генами (то есть миграции индивидуумов) между группами. Для того чтобы противодействовать отбору, поступление данного гена в субпопуляцию должно превышать его элиминацию отбором, а для того, чтобы противодействовать дрейфу, — превышать случайные потери этого гена. Когда такие субпопуляции более или менее изолированы, их называют демами. Демовая структура потенциально важна для эволюционных процессов, и мы обсудим этот вопрос в разд. 3.6. Генетический дрейф иногда называют эффектом Сьюэлла Райта и считают, что его действие направлено против действия отбора (рис. 2.9,Г). Однако, как это понимал Сьюэлл Райт, генетический дрейф может дать популяциям возможность покинуть локальные пики, пересечь имеющиеся в адаптивном ландшафте долины и подпасть под влияние факторов отбора, связанных с новыми пиками. Следовательно, генетический дрейф может способствовать адаптации, но может и препятствовать ей.
Дополнительно
Разработка сенсора на поверхностно-акустических волнах
В условиях
современности проблема контроля за состоянием окружающей среды выходит на все
более ведущее место. Контроль этот осуществляется как стационарными приборами, так и портативными. К стационарным приборам
можно отнести инфракрасные спектрометры, газовые хроматографы, массовые
спектрометры и ...
Порошковая металлургия и дальнейшая перспектива ее развития
Порошковой металлургией
называют область техники, охватывающую совокупность методов изготовления
порошков металлов и металлоподобных соединений, полуфабрикатов и изделий из них
или их смесей с неметаллическими порошками без расплавления основного
компонента.
Из имеющихся разнообразных
способов ...