Бактерии рода Clostridium
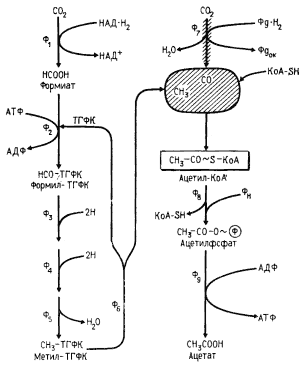
Вторая молекула CO2, углерод которой служит источником карбоксильной группы ацетата, при участии CO-дегидрогеназы восстанавливается до CO, оставаясь связанной с этим ферментом. CO-дегидрогеназа, ключевой фермент этого пути синтеза ацетата, имеет 3 центра, связывающие CH3-, CO-группы и кофермент А. Затем на ферменте осуществляется карбоксилирование метильной группы и взаимодействие образовавшейся ацетильной группы с коферментом А, приводящее к образованию ацетил-КоА. По характеру превращений, катализируемых CO-дегидрогеназой, фермент правильнее называть ацетил-КоА-синтазой. Дальнейшие превращения ацетил-КоА до ацетата, сопровождающиеся образованием АТФ, описаны ранее.
К протеолитическим относятся клостридии, имеющие активные протеолитические ферменты и поэтому способные использовать в качестве субстратов белки и пептиды, гидролизуя их до аминокислот и подвергая затем последние сбраживанию. В эту группу входят C. putrifiicum, C. histolyticum, C. sporogenes и другие сапрофитные виды. Близки к этим видам и некоторые патогенные формы: C. botulinum — продуцент ботулина — экзотоксина, являющегося одним из самых сильных биологических ядов; C. tetani — столбнячная палочка, образующая в организме человека столбнячный токсин. К протеолитическим клостридиям примыкают виды, использующие в качестве источника углерода и энергии ограниченное число свободных аминокислот. Например, C. cochlearium растет только на среде с глутаминовой кислотой, глутамином и гистидином; C. sticklandii может сбраживать лизин, аргинин, фенилаланин, серин, а C. propionicum — треонин, аланин, серин, цистеин.
|
|
|
Рис. 62. Ацетил-КоА-путь фиксации CO2 у гомоацетатных клостридиев: Ф1 — формиатдегидрогеназа; Ф2 — формил-ТГФК-синтаза; Ф3 — Ф5-ферменты, катализирующие превращение формил-ТГФК до метил-ТГФК; Ф6 — корриноидный фермент; Ф7 — CO-дегидрогеназа (фермент изображен в виде заштрихованных стрелки и эллипса); Ф8 — фосфотрансацетилаза; Ф9 — ацетаткиназа |
Известны два типа сбраживания аминокислот клостридиями. Для многих клостридиев одна аминокислота может служить источником энергии и углерода, например, глутаминовая кислота — для C. tetanomorphum, лизин — для C. sticklandii. В этом случае ее диссимиляция приводит к возникновению метаболитов, характерных для гликолитического пути, и в первую очередь пирувата, дальнейшие превращения которого идут по одному из путей, описанных выше. У C. sticklandii сбраживание лизина приводит к образованию масляной и уксусной кислот и NH3, а у C. tetanomorphum при сбраживании глутаминовой кислоты в дополнение к перечисленным выше продуктам образуется некоторое количество CO2.
Ряд аминокислот может подвергаться сбраживанию клостридиями только парами. Механизм процесса был расшифрован Л. Стиклендом (L. Stickland) в 1934 г., показавшим, что при этом происходит сопряженное окисление-восстановление пары аминокислот, одна из которых окисляется, другая — восстанавливается. Такой тип сбраживания аминокислот получил название реакции Стикленда. Окисляемыми аминокислотами, т. е. донорами электронов, служат аспарагин, аланин, валин, серин, гистидин и др. Восстанавливаемые аминокислоты — глицин, пролин, орнитин, аргинин и др.
Дополнительно
Галактика как уровень мегамира
Актуальность,
цели и задачи ответа по настоящей контрольной работе будут обусловлены
следующими положениями. Нас интересует не только звездное население того дома,
в котором мы живем. Нас интересует и архитектура этого дома и его размеры;
интересует, как его обитатели расселены, где жилищная тесно ...
Планета солнечной системы Уран
Даже в XVIII в.
планетная система была известна только до Сатурна. Но уже тогда предполагали,
что Сатурном список планет не оканчивается, что существуют еще более далекие
планеты, которые невооруженным глазом увидеть нельзя. Это мнение блестяще
подтвердилось, когда в 1781 г. знаменитый английский ...