Бактерии рода Clostridium
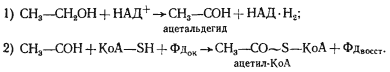
Электроны с ферредоксина могут переноситься далее на НАД+ или на Н+, что приводит в последнем случае к выделению H2. Следствием переноса части электронов на H+ будет нарушение соотношения между количеством ацетил-КоА и НАД-H2, необходимым для синтеза масляной кислоты (см. рис. 57), в сторону относительного недостатка молекул восстановленного кофермента. Возникшие "избыточные" молекулы ацетил-КоА используются для синтеза АТФ в реакциях, описанных ранее. По проведенным подсчетам, энергетический выход этого вида брожения составляет около 1 моля АТФ на 6 молей этанола.
Таким образом, типы брожений, осуществляемых клостридиями, необычайно разнообразны как в отношении используемых субстратов, так и синтезируемых конечных продуктов, и виды, осуществляющие сбраживание углеводов по гликолитическому пути с накоплением масляной кислоты в качестве одного из основных продуктов, являются только одной из групп организмов, относимых к роду Clostridium.
Особенности конструктивного метаболизма
Потребности клостридиев в питательных веществах отличаются большим разнообразием. Как правило, клостридии могут расти только на сложных, богатых органическими соединениями средах. Многие клостридии выделяют экзоферменты, расщепляющие макромолекулы (углеводы, белки) на составляющие их мономеры. До сих пор только небольшое число видов удалось культивировать в лаборатории на синтетической среде. Для них выявлена потребность в витаминах (главным образом группы В) и наборе аминокислот.
Интересная особенность эубактерий из рода Clostridium — дальнейшее развитие способности вовлекать углекислоту в клеточный метаболизм. У C. kluyveri, растущего на смеси C3-соединений (этанол + ацетат), до 30% углерода клетки возникает из углерода CO2. Для C. aceticum и других видов, осуществляющих гомоацетатное брожение, показана способность к хемолитоавтотрофному росту на минеральной среде за счет превращения CO2 и H2 в ацетат:
4H2 + 2CO2 ® CH3-COOH + 2H2O.
В группу гомоацетатных бактерий, помимо некоторых представителей рода Clostridium, входят организмы, принадлежащие к родам Acetobacterium, Acetogenium, Eubacterium, Butyribacterium и др. При росте на минеральной среде энергию ацетогены получают в процессе анаэробного дыхания с использованием H2 в качестве донора электронов, а CO2 — конечного их акцептора (см. гл. 16). Исходными метаболитами для биосинтетических путей служат ацетил-КоА (ключевой метаболит пути фиксации CO2 ацетогенами) и пируват, образующийся из последнего в реакции восстановительного карбоксилирования.
Пути включения CO2 в клеточный метаболизм клостридиев различны. Углекислота может использоваться ими в качестве конечного акцептора электронов, что приводит к прямому восстановлению CO2 до формиата. Донорами электронов в этой реакции служат восстановленный ферредоксин или НАД-H2. Реакция может служить способом удаления избытка восстановительных эквивалентов, образующихся при брожении, т. е. быть необходимой для сбалансирования окислительных и восстановительных этапов в энергетическом метаболизме. Образовавшийся в результате восстановления CO2 формиат может подвергаться дальнейшему восстановлению и служить источником метильных групп, используемых для клеточных биосинтезов.
Дополнительно
Эвристика и ее применение
В своей повседневной жизни человек все время
сталкивается с задачами легкими для него, но с трудом решаемыми машинами.
Тяжело создать программу, которая предусматривала бы все. Поэтому в условиях
недостаточности или сложности информации человек практически незаменим.
Преодолеть же пропасть между м ...
Автоматизированное проектирование станочной оснастки
1.1.
СТАНОЧНЫЕ ПРИСПОСОБЛЕНИЯ .
КЛАССИФИКАЦИЯ , ВИДЫ .
1.1.1. Станочные
приспособления .
Основную группу
технологической оснастки составляют приспособления механосборочного
производства. Приспособлениями в машиностроении называют вспомогательные
устройства к технологич ...