Структура, химический состав и функции компонентов прокариотной клетки
Изучение строения жгутика под электронным микроскопом обнаружило, что он состоит из трех частей (рис. 11). Основную массу жгутика составляет длинная спиральная нить (фибрилла), у поверхности клеточной стенки переходящая в утолщенную изогнутую структуру—крюк. Нить с помощью крюка прикреплена к базальному телу, вмонтированному в ЦПМ и клеточную стенку. У большинства прокариот нить состоит только из одного типа белка — флагеллина. Белковые субъединицы уложены в виде спирали, внутри которой проходит полый канал. Наращивание жгутика происходит с дистального конца, куда субъединицы поступают по внутреннему каналу. У некоторых видов жгутик снаружи дополнительно покрыт чехлом особого химического строения или же являющимся продолжением клеточной стенки и, вероятно, построенным из того же материала.
Крюк (толщина 20—45 нм) состоит из белка, отличающегося от флагеллина, и служит для обеспечения гибкого соединения нити с базальным телом. Базальное тело содержит 9—12 различных белков и представляет собой систему из двух или четырех колец, нанизанных на стержень, являющийся продолжением крюка. Два внутренних кольца (M и S) — обязательные составные части базального тела, в то время как наружные кольца (Р и L) отсутствуют у грамположительных эубактерий и, следовательно, не необходимы для движения. M-кольцо локализовано в ЦПМ, S-кольцо располагается в периплазматическом пространстве грамотрицательных или в пептидогликановом мешке грамположительных эубактерий.
|
|
|
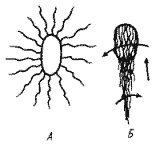
Рис. 12. Клетка Salmonella typhimurium в состоянии покоя (А) и при движении (Б). Стрелками показано направление вращения и движения клетки |
Кольца Р и L, имеющиеся только у грамотрицательных эубактерий, локализованы соответственно в пептидогликановом слое и в наружной мембране. Особенности строения базального тела определяются, таким образом, строением клеточной стенки. Интактность последней необходима для движения жгутиковых бактерий. Обработка клеток лизоцимом, приводящая к удалению пептидогликанового слоя клеточной стенки, вызывает и потерю способности бактерий к движению, хотя жгутики остаются при этом неповрежденными.
Предполагают, что вращение жгутика определяется вращением M-кольца. Другие кольца базального тела неподвижны и служат для крепления стержня, проходящего через клеточную стенку грамотрицательных эубактерий. У грамположительных эубактерий эту функцию в основном выполняет многослойный жесткий пептидогликановый мешок.
Большие успехи достигнуты в расшифровке механизма движения прокариот, имеющих жгутики. Если в клетке много жгутиков. все они при движении собираются в пучок, вращаясь в одном направлении (рис. 12). Вращение жгутиков передается клетке, начинающей вращаться в противоположном направлении, и обеспечивает эффективное движение (плавание) в жидкой среде и более медленное перемещение по поверхности твердых сред.
Для работы двигательного аппарата прокариот необходима энергия. Установлено, что движение жгутиковых прокариот обеспечивается энергией трансмембранного электрохимического потенциала (DmH+), причем обе его составляющие — электрическая (Dy) и концентрационная (DpH) — поддерживают движение. Скорость вращения жгутиков прямо зависит от величины мембранного потенциала. Таким образом, прокариотная клетка обладает механизмом, позволяющим превращать электрохимическую форму энергии непосредственно в механическую. Молекулярное устройство, обеспечивающее это превращение, к настоящему времени не выяснено, но можно полагать, что оно должно быть весьма эффективным, так как, по проведенным расчетам, энергия, расходуемая на движение, составляет десятые доли процента от общего количества энергетических потребностей клетки.
|
|
|
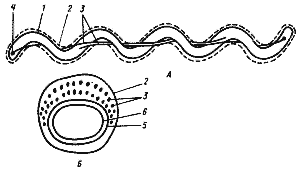
Рис. 13. Клетка спирохеты в продольном (А) и поперечном (Б) разрезе. На рис. А изображена клетка, содержащая по одной аксиальной фибрилле у каждого конца; на рис. Б — поперечный разрез, прошедший через среднюю часть клетки, где показаны два пересекающихся пучка, состоящих из множества аксиальных фибрилл: 1 — протоплазматический цилиндр; 2 — наружный чехол; 3 — аксиальные фибриллы; 4 — место прикрепления аксиальных фибрилл; 5 — пептидогликановый слой клеточной стенки; 6 — ЦПМ |
Дополнительно
Современная судовая газотурбинная установка
Современная
судовая газотурбинная установка (ГТУ) успешно конкурирует с аналогичными по
назначению паротурбинными и дизельными. От последних она выгодно отличается
компактностью и малой удельной массой, маневренностью и высокой
ремонтопригодностью, лучшей приспособленностью к автоматизации ...
Расчет релаксационного генератора на ИОУ
Разработать и рассчитать
релаксационный генератор на ИОУ
(интегральной схеме операционного
усилителя) в соответствии с данными, представленными:
·
вид генератора - мультивибратор
·
режим работы – автоколебательный
·
период следования импульсов Т, мс – 0.09
· ...